马дёҠжіЁеҶҢпјҢз»“дәӨжӣҙеӨҡеҘҪеҸӢпјҢдә«з”ЁжӣҙеӨҡеҠҹиғҪпјҢи®©дҪ иҪ»жқҫзҺ©иҪ¬зӨҫеҢәгҖӮ
жӮЁйңҖиҰҒ зҷ»еҪ• жүҚеҸҜд»ҘдёӢиҪҪжҲ–жҹҘзңӢпјҢжІЎжңүиҙҰеҸ·пјҹз«ӢеҚіжіЁеҶҢ

x
дәәзұ»иӮ йҒ“еҗ«жңүдё°еҜҢзҡ„з»ҶиҸҢпјҢе…¶ж•°йҮҸжҳҜдәәдҪ“з»Ҷиғһж•°йҮҸзҡ„10еҖҚпјҢд»ҺдёҠеҫҖдёӢз»ҶиҸҢи¶ҠжқҘи¶ҠеӨҡпјҡеҚҒдәҢжҢҮиӮ 102~103/mlпјҢз©әиӮ 103~105/mlпјҢеӣһиӮ 105~107/mlпјҢеӨ§иӮ 1010~1012/mlгҖӮиӮ йҒ“иҸҢзҫӨпјҲgut microbiotaпјүзҡ„йҮҚйҮҸзәҰеҚ дәәдҪ“дҪ“йҮҚзҡ„2зЈ…е·ҰеҸіпјҢе…¶еҹәеӣ йҮҸзәҰ3зҷҫдёҮдёӘеҹәеӣ пјҢжҳҜдәәдҪ“еҹәеӣ йҮҸзҡ„100еҖҚгҖӮиӮ йҒ“иҸҢзҫӨзҡ„з»ҶиҸҢз»„жҲҗпјҢеңЁй—Ёзә§пјҲphylum levelпјүж°ҙе№ідёҠпјҢеҺҡеЈҒиҸҢй—ЁпјҲFirmicutesпјүеҸҠжӢҹжқҶиҸҢй—ЁпјҲBacteriodetesпјүзәҰеҚ 90%пјҢе…¶дҪҷдёәи“қи—»з»ҶиҸҢпјҲCyanobacteriaпјүпјҢеҸҳеҪўиҸҢй—ЁпјҲProteobacteriaпјүпјҢж”ҫзәҝиҸҢпјҲActinobacteriaпјүпјҢжўӯиҸҢеұһпјҲFusobacteriaпјүеҸҠз–Јеҫ®иҸҢй—ЁпјҲVerrumicrobioa phylaпјүгҖӮз»ҸиҝҮ5дәҝе№ҙзҡ„иҝӣеҢ–пјҢиҝҷдәӣеҫ®з”ҹзү©дёҺе®ҝдё»е·Із»ҸеҪўжҲҗдәҶдёҖдёӘе‘Ҫиҝҗе…ұеҗҢдҪ“пјҢжҲҗдёәе®ҝдё»зҡ„еҸҰеӨ–дёҖдёӘйҮҚиҰҒеҷЁе®ҳпјҢиҝҷдәӣе…ұз”ҹиҸҢзҫӨпјҲcommensal microbiotaпјүйҖҡиҝҮз”ҹзү©жҙ»жҖ§еҲҶеӯҗдёҺе®ҝдё»дәӨжөҒпјҢеҪұе“Қе®ҝдё»зҡ„еӨҡз§Қе…Қз–«еҸҠйқһе…Қз–«еҠҹиғҪ[1]пјҢи§Ғеӣҫ1гҖӮ
еӣҫзүҮиҜ·зӮ№еҮ»дёҠж–№й“ҫжҺҘ
1 иӮ йҒ“иҸҢзҫӨи°ғжҺ§е…Қз–«еҸҚеә”
з”ҹе‘Ҫж—©е№ҙзҡ„еҫ®з”ҹзү©жҡҙйңІеҜ№еҠЁзү©ж—ҘеҗҺжңүй•ҝжңҹзҡ„еҪұе“ҚпјҢе…Қз–«зі»з»ҹзҡ„е……еҲҶеҸ‘иӮІжңүиө–дәҺеҮәз”ҹеҗҺе…ұз”ҹиҸҢзҫӨзҡ„жҡҙйңІдёҺжҺҘи§ҰпјҢиӮ йҒ“зӣёе…іжҖ§ж·Ӣе·ҙз»„з»ҮпјҲgut-associ-ated lymphoid tissues, GALTsпјүе®Ңе…ЁжҲҗзҶҹгҖҒдә§IgAжөҶз»Ҷиғһе’ҢжҝҖжҙ»Tз»ҶиғһеӢҹйӣҶеҲ°й»ҸиҶңпјҢеқҮжңүиө–дәҺеҮәз”ҹеҗҺиҸҢзҫӨй©ұеҠЁдҝЎеҸ·еҜ№дёҠзҡ®з»ҶиғһеҸҠж ‘зӘҒзҠ¶з»ҶиғһпјҲdendritic cells, DCsпјүзҡ„дҪңз”ЁгҖӮ延иҝҹз»ҶиҸҢжҡҙйңІзҡ„ж— иҸҢпјҲgerm-free, GFпјүе°Ҹйј пјҢзӮҺз—ҮжҖ§иӮ з—…пјҲin?ammatory bowel disease, IBDпјүеҸҠиҝҮж•ҸжҖ§е“®е–ҳжӯ»дәЎзҺҮжӣҙй«ҳ[2]гҖӮ
е®ҡжӨҚеңЁиӮ йҒ“й»ҸиҶңдёҠзҡ®иЎЁйқўзҡ„иҸҢзҫӨеҸҜд»ҘзӣҙжҺҘдҪңз”ЁдәҺе®ҝдё»жҲ–иҖ…йҖҡиҝҮдә§зү©еҰӮиӣӢзҷҪиҙЁгҖҒи„ӮиҙЁгҖҒзўіж°ҙеҢ–еҗҲзү©еҸҠж ёй…ёй—ҙжҺҘдҪңз”ЁдәҺе®ҝдё»пјҢи°ғиҠӮеұҖйғЁзӮҺз—ҮеҸҠе…Қз–«гҖӮе®ҝдё»дёҺиӮ йҒ“иҸҢзҫӨзҡ„з”ҹзҗҶжҖ§зӣёдә’дҪңз”ЁеҸҜд»ҘзӣҙжҺҘжҠөеҫЎе…ұз”ҹиҸҢеҰӮе°ҸиӮ д№ій…ёжқҶиҸҢпјҲLactobacilliпјүе’ҢеҸҳеҪўиҸҢпјҲProteobacteriaпјүгҖҒз»“иӮ жўӯзҠ¶иҠҪиғһжқҶиҸҢпјҲClostridiaпјүе’ҢжӢҹжқҶиҸҢпјҲBacteroidesпјүпјҢйҒҝе…Қз—…еҺҹиҸҢеҰӮжІҷй—Ёж°ҸиҸҢпјҲSalmonellaпјүе’Ңеҝ—иҙәж°ҸжқҶиҸҢпјҲShigella spp.пјүж„ҹжҹ“пјҢжҺ§еҲ¶еҶ…жәҗжҖ§з—…зҗҶз”ҹзү©пјҲpathobiontsпјүеҰӮиү°йҡҫжўӯиҸҢпјҲClostridium diffcileпјүе’ҢиӮ зҗғиҸҢпјҲEnterococciпјүзҡ„иҝҮеәҰз№Ғж®–[3]гҖӮ
е…ұз”ҹиҸҢзҫӨдёҚд»…и°ғиҠӮиӮ йҒ“еұҖйғЁе…Қз–«пјҢиҖҢдё”и°ғиҠӮе…Ёиә«е…ҲеӨ©жҖ§е…Қз–«еҸҠиҺ·еҫ—пјҲеҗҺеӨ©пјүжҖ§е…Қз–«гҖӮзҡ®иӮӨгҖҒйҳҙйҒ“гҖҒеҸЈи…”зӯүеұҸйҡңиЎЁйқўзҡ„иҸҢзҫӨйҖҡеёёеҸӘеҜ№еұҖйғЁе…Қз–«иҙҹиҙЈпјҢдҪҶжҳҜпјҢиӮ йҒ“иҸҢзҫӨеҲҷе…·жңүе…Ёиә«жҖ§е…Қз–«иҮӘзЁідҪңз”ЁпјҢе…¶жңәеҲ¶еҸҜиғҪдёҺиӮ йҒ“иҸҢзҫӨз§Қзұ»з№ҒеӨҡгҖҒж•°йҮҸе·ЁеӨ§гҖҒиӮ йҒ“й»ҸиҶңйқўз§Ҝе№ҝйҳ”жңүе…і[4]гҖӮиӮ йҒ“зјәд№Ҹе…ұз”ҹиҸҢзҫӨзҡ„е°Ҹйј пјҢзӮҺз—ҮеҸҚеә”еҸҠе…Қз–«еә”зӯ”иғҪеҠӣдёӢйҷҚпјҢеҜ№еҗ„з§ҚеҲәжҝҖдёҚиғҪеҪўжҲҗзҗҶжғізҡ„йҳІеҫЎеҸҚеә”пјҢдёҚиғҪжңүж•ҲжҠөжҠ—еӨҡдёӘи§Јеү–йғЁдҪҚзҡ„ж„ҹжҹ“пјҢеҰӮиӮәз”ІеһӢH1N1жөҒж„ҹгҖҒеҸЈи…”жқҺж–Ҝзү№иҸҢж„ҹжҹ“пјҢиҖҢдё”йҡҫд»ҘиҜұеҜјиҮӘиә«е…Қз–«жҖ§з–ҫз—…еҠЁзү©жЁЎеһӢеҰӮеӨҡеҸ‘жҖ§зЎ¬еҢ–гҖҒе…іиҠӮзӮҺпјҢз”ҡиҮіеҜ№еӨҡз§ҚжҠ—иӮҝзҳӨе…Қз–«жІ»з–—еҸҠеҢ–з–—дёҚж•Ҹж„ҹ[5]гҖӮи§ӮеҜҹеҸ‘зҺ°[6] пјҡGF е°Ҹйј гҖҒжҠ—з”ҹзҙ жІ»з–—е°Ҹйј еӨҡдёӘдәҡзҫӨзҡ„Tз»ҶиғһеҰӮTh1гҖҒTh17пјҲдәҢиҖ…еҲҶеҲ«дә§з”ҹIFN-Оіе’ҢIL-17пјүж•°йҮҸдёӢйҷҚпјҢTregз»Ҷиғһж•°йҮҸеўһеҠ пјҢе…Ёиә«ж„ҹжҹ“дёҚиғҪжҝҖеҸ‘иҝҷдәӣе°Ҹйј зҡ„е…ҲеӨ©жҖ§е’ҢеҗҺеӨ©жҖ§е…Қз–«еҸҚеә”пјҢи§Ғеӣҫ2гҖӮ
еӣҫзүҮиҜ·зӮ№еҮ»дёҠж–№й“ҫжҺҘ
д»ҘеүҚдәә们дёҖзӣҙи®Өдёә пјҡиӮ йҒ“иҸҢзҫӨеҜ№дәәдҪ“е…Қз–«жҲҗзҶҹзҡ„еҲәжҝҖдҪңз”Ёдҫқиө–ж•ҙдёӘиҸҢзҫӨзҡ„е…ұеҗҢдҪңз”ЁгҖӮзҺ°еңЁеҸ‘зҺ°пјҢиӮ йҒ“зҡ„еҚ•дёӘз»ҶиҸҢеҚіеҸҜд»ҘеҲәжҝҖе…Қз–«еә”зӯ”гҖӮеҲҶиҠӮдёқзҠ¶иҸҢпјҲsegmented flamentous bacteria, SFBпјүжҳҜдёҖз§ҚдёҚеҸҜеҹ№е…»зҡ„з»ҶиҸҢпјҢеҸҜд»ҘеҲәжҝҖе°Ҹйј й»ҸиҶңе…Қз–«еә”зӯ”зҡ„жҲҗзҶҹгҖӮзјәд№Ҹ SFBпјҢе°Ҹйј IgA ж»ҙеәҰдёӢйҷҚгҖҒй»ҸиҶңTh1еҸҠTh17з»Ҷиғһж°ҙе№ійҷҚдҪҺпјҢеҜ№иӮ йҒ“иҮҙз—…иҸҢCitrobacter rodentiumгҖҒSalmonella spp.зҡ„еҸҚеә”дёҚиүҜ[7]гҖӮ
Bacteroides fragilisйҖҡиҝҮи°ғиҠӮTLR2жҝҖеҠЁеүӮвҖ”еӣҠеӨҡзі–пјҲcapsular polysaccharideпјүAй©ұеҠЁдә§IL-10 Tregз»Ҷиғһзҡ„еҲҶеҢ–пјҢдҝқжҠӨе°Ҹйј е…ҚеҸ—иӮқиһәжқҶиҸҢпјҲHelicobacter hepaticusпјүж„ҹжҹ“д»ҘеҸҠдёүзЎқеҹәиӢҜзЈәй…ёпјҲtrinitrobenzene sulfonic acid, TNBSпјүиҜұеҜјзҡ„з»“иӮ зӮҺгҖӮ
2 иӮ йҒ“иҸҢзҫӨи°ғиҠӮиғҪйҮҸд»Ји°ў
дәәдҪ“е°ҶеӨҡдҪҷиғҪйҮҸеӮЁеӯҳдёәи„ӮиӮӘд»Ҙи°ғиҠӮдәәдҪ“иғҪйҮҸе№іиЎЎпјҢиӮ йҒ“иҸҢзҫӨеҸӮдёҺдәҶиҝҷдёҖиҝҮзЁӢ[8]гҖӮе®һйӘҢжҳҫзӨәпјҡе°Ҫз®ЎGFе°Ҹйј жҜ”жҷ®йҖҡе°Ҹйј жҜҸеӨ©еӨҡеҗғ30%зҡ„йЈҹзү©пјҢдҪҶжҳҜGFе°Ҹйј зҡ„жҖ»дҪ“и„ӮжҜ”жҷ®йҖҡе°Ҹйј е°‘40%гҖӮжҠҠз»ҶиҸҢе®ҡжӨҚе…ҘGFе°Ҹйј иӮ йҒ“еҶ…пјҢе°ҶGFйј еҸҳдёәжңүиҸҢйј пјҲиҪ¬еҢ–йј пјҢдёӢеҗҢпјүпјҢе°Ҹйј жҜҸеӨ©ж‘„йЈҹйҮҸе°Ҫз®ЎжңүжҳҺжҳҫеҮҸе°‘пјҢдҪҶжҳҜеңЁдёӨе‘ЁеҶ…е°Ҹйј дҪ“йҮҚеҚҙжҳҺжҳҫеўһеҠ пјҢдҪ“еҶ…и„ӮиӮӘеўһеҠ 60%пјҢ并且еҸ‘з”ҹиғ°еІӣзҙ жҠөжҠ—[9]гҖӮиҝҷз§ҚдҪ“йҮҚзҡ„еўһеҠ жҳҫ然жҳҜз”ұдәҺиӮ йҒ“иҸҢзҫӨе°Ҷе®ҝдё»ж‘„е…Ҙзҡ„дёҚж¶ҲеҢ–еӨҡзі–еҰӮжҠ—жҖ§ж·ҖзІүгҖҒеҜЎиҒҡзі–гҖҒиҸҠзІүиҝӣиЎҢеҸ‘й…өйҷҚи§ЈпјҢиӮ йҒ“и‘Ўиҗ„зі–ж‘„еҸ–еўһеҠ жүҖиҮҙпјҢи§Ғеӣҫ3гҖӮ
еӣҫзүҮиҜ·зӮ№еҮ»дёҠж–№й“ҫжҺҘ
иӮ йҒ“иҸҢзҫӨдёҺиӮҘиғ–зҡ„е…ізі»йҰ–е…ҲеңЁе•®йҪҝеҠЁзү©жЁЎеһӢдёҠеҫ—еҲ°иҜҒе®һгҖӮLey REзӯү[10]и§ӮеҜҹдәҶйҒ—дј жҖ§иӮҘиғ– ob/obе°Ҹйј е’ҢеҜ№еә”зҳҰйј зҡ„5000еӨҡдёӘиӮ йҒ“иҸҢзҫӨ16S RNAеҹәеӣ еәҸеҲ—пјҢеҸ‘зҺ°ob/obиӮҘйј зҡ„жӢҹжқҶиҸҢй—ЁеҮҸе°‘50%пјҢзЎ¬еЈҒиҸҢй—ЁжңүеҗҢж ·жҜ”дҫӢзҡ„еўһеҠ гҖӮдәәзұ»з ”究дёӯи§ӮеҜҹ12еҗҚиӮҘиғ–еҸ—иҜ•иҖ…еҸ‘зҺ°пјҢдёҺж¶ҲзҳҰеҸ—иҜ•иҖ…жҜ”иҫғпјҢйҘ®йЈҹе№Ійў„еүҚиӮҘиғ–еҸ—иҜ•иҖ…иӮ йҒ“жӢҹжқҶиҸҢй—ЁиҫғдҪҺпјҲеҮҸе°‘90%пјүпјҢзЎ¬еЈҒиҸҢй—Ёиҫғй«ҳпјҲеўһеӨҡ20%пјүпјҢз»ҸиҝҮ52е‘ЁйҷҗеҲ¶йҘ®йЈҹд№ӢеҗҺпјҢеҸ—иҜ•иҖ…дҪ“йҮҚдёӢйҷҚпјҢиӮ йҒ“жӢҹжқҶиҸҢй—Ё/зЎ¬еЈҒиҸҢй—ЁжҜ”дҫӢжҺҘиҝ‘зҳҰдәәгҖӮ Turnbaugh PJзӯү[11]еҲҶеҲ«е°Ҷob/obиӮҘйј еҸҠзҳҰйј еӣһиӮ з»ҶиҸҢ移жӨҚеҲ°йҮҺз”ҹеһӢGFиӮ йҒ“пјҢдёҺ移жӨҚзҳҰйј иӮ йҒ“иҸҢзҫӨзҡ„GFе°Ҹйј зӣёжҜ”пјҢ2е‘ЁеҗҺ移жӨҚиӮҘйј иӮ йҒ“иҸҢзҫӨзҡ„GFйј е…¶дҪ“и„Ӯеҗ«йҮҸжңүдёӯзӯүеўһеҠ пјҢд»ҺйЈҹзү©дёӯж‘„еҸ–иғҪйҮҸжӣҙеӨҡгҖӮиҜҙжҳҺиӮ йҒ“иҸҢзҫӨзӣёе…іжҖ§жңәеҲ¶еңЁйҘ®йЈҹзӣёе…іжҖ§д»Ји°ўзҙҠд№ұгҖҒиӮҘиғ–зҡ„еҸ‘з”ҹеҸ‘еұ•иҝҮзЁӢдёӯжү®жј”йҮҚиҰҒи§’иүІпјҢжҸҗзӨәиӮ йҒ“иҸҢзҫӨеҸӮдёҺе®ҝдё»д»Ји°ўжҖ§з–ҫз—…зҡ„еҸ‘з”ҹ[12]гҖӮ
3 е…ұз”ҹиҸҢзҫӨеҸҠиҸҢзҫӨеӨұи°ғеңЁиӮҝзҳӨеҸ‘з”ҹдёӯзҡ„дҪңз”Ё
з”ҹзҗҶжқЎд»¶дёӢпјҢиӮ йҒ“еҫ®з”ҹзү©зҫӨдёҚд»…дёҚжҳҜиҮҙзҷҢеӣ зҙ пјҢеҸҚиҖҢжҳҜжҠ—зҷҢеӣ зҙ гҖӮиӮ йҒ“иҸҢзҫӨе…·жңүж—¶й—ҙзЁіе®ҡжҖ§
е’Ңеӣһеј№жҖ§пјҢйҒҮеҲ°жҚҹдјӨеҸҜд»ҘиҮӘе·ұдҝ®еӨҚ[13]гҖӮдҪҶжҳҜеҰӮжһңиӮ йҒ“иҸҢзҫӨз»„жҲҗзҡ„еҸҳеҢ–и¶…иҝҮдәҶе…¶еӣһеј№иғҪеҠӣпјҢеҲҷдјҡеј•иө·иҸҢзҫӨеӨұи°ғпјҲdysbiosisпјүгҖӮиҸҢзҫӨеӨұи°ғеҜјиҮҙз—…еҺҹеҫ®з”ҹзү©жҺ§еҲ¶еӨұиҙҘгҖҒзӮҺз—ҮжҲ–е…Қз–«еҸҚеә”еӨұи°ғпјҢеј•иө·жҖҘжҖ§жҲ–ж…ўжҖ§з»„з»ҮжҚҹдјӨгҖӮ
иӮ йҒ“еұҸйҡңеҠҹиғҪз ҙеқҸеҗҺзҡ„иӮ йҒ“иҸҢзҫӨеӨұи°ғжң¬иә«еҸҠе…¶дёҺйҖ иЎҖз»Ҷиғһзҡ„зӣёдә’дҪңз”Ёи°ғиҠӮзӮҺз—ҮеҸҚеә”пјҢжҲҗдёәиӮҝзҳӨзҡ„еҸ‘з—…еҺҹеӣ гҖӮ IL-18е…·жңүй»ҸиҶңдҝқжҠӨдҪңз”ЁгҖӮзјәIL-18гҖҒIL-18RгҖҒMyD88гҖҒзӮҺз—Үз»„пјҲin?ammasomesпјүеҸҠзӮҺз—Үз»„дҝЎеҸ·еҲҶеӯҗзҡ„е°Ҹйј пјҢдёҚиғҪдә§з”ҹIL-18гҖҒдёҚиғҪеӨ„зҗҶпјҲprocessпјүIL-18гҖҒеҜ№IL-18д№ҹдёҚиғҪеҸҚеә”пјҢиҝҷдәӣе°Ҹйј зҡ„иӮ йҒ“иҸҢзҫӨеӨұи°ғпјҢBacteroidetesпјҲPrevotellaceaeпјүеҸҠTM7жҜ”дҫӢеҚҮй«ҳпјҢеҢ–еӯҰжҲҗзҳӨж•Ҹж„ҹ[16]гҖӮе°Ҷиҝҷдәӣе°Ҹйј дёҺеҒҘеә·е°Ҹйј е…ұеұ…пјҲcohousingпјүпјҢжҲ–иҖ…е°Ҷе®ғ们зҡ„зІӘдҫҝ移жӨҚз»ҷеҒҘеә·е°Ҹйј пјҢеҒҘеә·е°Ҹйј е®№жҳ“еҸ‘з”ҹз»“иӮ зҷҢ[14]гҖӮ
дёҖдәӣзү№е®ҡзҡ„з»ҶиҸҢеҰӮStreptococcus gallolyticusпјҢEnterococcus faecalis, enterotoxigenic B. Fragilis, enteropathogenic Escherichia coliеҸҠFusobacterium spp еҸҜд»ҘиҜұеҜјз»“иӮ зҷҢ[15]гҖӮз»“иӮ зҷҢиҚ·зҳӨе°Ҹйј пјҢ Odoribacterе’ҢAkkermansiaеҹәеӣ еҜҢйӣҶгҖӮдёҖдәӣзІӘиҸҢеҰӮMethanobacterialesдёҺз»“иӮ зҷҢзҡ„еҸ‘з”ҹеҜҶеҲҮзӣёе…іпјҢFusobacterium nucleatumйҖҡиҝҮдҝғиҝӣFadAдёҺE-cadherinзҡ„зІҳеҗҲпјҢжҝҖжҙ»ОІ-catenin/WntдҝЎеҸ·пјҢиҜұеҜјдҝғзҷҢй«“зі»з»Ҷиғһзҡ„еўһж®–дёҺжҙ»еҢ–гҖӮдёҠиҝ°з»ҶиҸҢдёҚд»…еҸҜд»ҘиҮҙзҷҢпјҢиҝҳеҸҜд»ҘйҖҡиҝҮеӨҡз§ҚжңәеҲ¶еҰӮз»ҶиҸҢзҫӨдҪ“ж„ҹеә”пјҲquorum sensingпјүзҡ„дҝЎжҒҜдәӨжөҒгҖҒжҝҖзҙ еҸҠжҠ—иҸҢеӣ еӯҗзҡ„еҲҶжіҢпјҢе…ұеҗҢдҪңз”Ёж”№еҸҳиӮ йҒ“иҸҢзҫӨзҡ„з»„жҲҗпјҢеј•иө·иҸҢзҫӨеӨұи°ғ[15]гҖӮз ”з©¶еҸ‘зҺ°пјҡдёҺе‘ЁеӣҙжӯЈеёёз»„з»ҮзӣёжҜ”пјҢиӮ йҒ“жҒҜиӮүеҜ№з»ҶиҸҢеҫ—йҖҡйҖҸжҖ§жӣҙй«ҳгҖҒжӣҙе®№жҳ“еҸ‘з”ҹз»ҶиҸҢ移дҪҚгҖӮиҝҷдәӣ移дҪҚзҡ„з»ҶиҸҢйҖҡиҝҮиҜұеҜјдҝғзҷҢз»Ҷиғһеӣ еӯҗIL-6гҖҒIL-11гҖҒIL-23гҖҒIL-17гҖҒIL-22зҡ„дә§з”ҹпјҢиҜұеҜјзӮҺз—ҮеҸҚеә”пјҢдҝғиҝӣжҒҜиӮүиҝӣеұ•[16]гҖӮ
иӮ йҒ“иҸҢзҫӨдёӯзҡ„дёҖдәӣз»ҶиҸҢдёҚд»…е…·жңүеұҖйғЁиҮҙзҷҢдҪңз”ЁпјҢиҝҳеҸҜд»ҘиҜұеҜјиҝңйҡ”йғЁдҪҚзҡ„иӮҝзҳӨгҖӮз»“иӮ H. hepaticusж„ҹжҹ“дёҚд»…еҸҜд»ҘеҚҮй«ҳAPCmin/+е°Ҹйј гҖҒж°§еҢ–еҒ¶ж°®з”Ізғ·еӨ„зҗҶRag2?/? е°Ҹйј зҡ„е°ҸиӮ еҸҠз»“иӮ зҡ„иӮҝзҳӨеҸ‘з”ҹзҺҮпјҢиҖҢдё”еҚҮй«ҳдәҶAPCmin/+/Rag2?/?е°Ҹйј зҡ„д№іи…әзҷҢеҸҠеүҚеҲ—и…әзҷҢзҡ„еҸ‘з—…зҺҮпјҢиҝҳжҸҗй«ҳдәҶе°Ҹйј иӮқи„ҸеҜ№еҢ–еӯҰиҮҙзҷҢзү©гҖҒз—…жҜ’иҮҙзҷҢзҡ„ж•Ҹж„ҹжҖ§[17]гҖӮиӮ йҒ“иҸҢзҫӨиҜұеҜјиҝңйҡ”йғЁдҪҚиӮҝзҳӨзҡ„жңәеҲ¶иҝҳдёҚжё…жҘҡпјҢеҸҜиғҪйғЁеҲҶдёҺи°ғиҠӮ TNFдҫқиө–зҡ„е…Ёиә«зӮҺз—ҮеҸҚеә”еј еҠӣпјҲsystemic in?ammatory toneпјүгҖҒж°§еҢ–еә”жҝҖгҖҒзҷҪз»ҶиғһеҸҠдёҠзҡ®з»Ҷиғһзҡ„еҹәеӣ жҜ’жҖ§пјҲgenotoxicityпјүжңүе…ігҖӮиҸҢзҫӨеӨұи°ғеҸҠжҠ—з”ҹзҙ жІ»з–—еҸҜд»Ҙж”№еҸҳиӮ йҒ“иҸҢзҫӨеҜ№йӣҢжҝҖзҙ зҡ„д»Ји°ўиғҪеҠӣпјҢиҜҘиғҪеҠӣиў«и®ӨдёәжҳҜиӮ йҒ“иҸҢзҫӨи°ғиҠӮиҝңйҡ”йғЁдҪҚиӮҝзҳӨзҡ„жңәеҲ¶д№ӢдёҖ[18]гҖӮеҸҰдёҖж–№йқўпјҢиӮ йҒ“иҸҢзҫӨд№ҹе…·жңүжҠ—иӮҝзҳӨдҪңз”ЁгҖӮиӮ йҒ“иҸҢзҫӨеңЁз»“иӮ еҸ‘й…өиҶійЈҹзәӨз»ҙгҖҒдә§з”ҹзҹӯй“ҫи„ӮиӮӘй…ёпјҲshort chain fatty acids, SCFAsпјү пјҢ еҗҺиҖ…йҖҡиҝҮв‘ жҝҖжҙ»GиӣӢзҷҪеҒ¶иҒ”еҸ—дҪ“пјҲG-protein coupled receptors, GPRпјү109aпјҢиҜұеҜјиӮ йҒ“дёҠзҡ®з»ҶиғһеҲҶжіҢIL-18пјӣв‘ЎзӣҙжҺҘдҪңз”ЁдәҺDCгҖҒе·Ёеҷ¬з»ҶиғһеҸҠTз»Ҷиғһпјӣв‘ўиҜұеҜјTregз»Ҷиғһеўһж®–пјҢзӣҙжҺҘи°ғиҠӮTregз»Ҷиғһжұ зҡ„еӨ§е°Ҹе’ҢеҠҹиғҪпјҢдә§з”ҹжҠ—зӮҺз»Ҷиғһеӣ еӯҗIL-10пјҢд»ҺиҖҢжҠ‘еҲ¶з»“иӮ зӮҺз—ҮеҸҚеә”гҖҒжҠ‘еҲ¶иӮҝзҳӨеҪўжҲҗ[19] гҖӮIL-18йҖҡиҝҮи°ғиҠӮIL-22зҡ„дә§з”ҹе’ҢдҪңз”ЁпјҢдҝғиҝӣиӮ йҒ“й»ҸиҶңзҡ„дҝ®еӨҚгҖӮIL-22з”ұиӮ йҒ“й»ҸиҶңеӣәжңүеұӮзҡ„еҲқе§Ӣж·Ӣе·ҙз»Ҷиғһдә§з”ҹгҖҒйҖҡиҝҮжҝҖжҙ»STAT3пјҢиҜұеҜјдёҠзҡ®з»Ҷиғһеўһ殖并дә§з”ҹжҠ—иҸҢиӮҪпјҢеӣ жӯӨеҸ‘жҢҘжҠ‘зҷҢеҪўжҲҗдҪңз”ЁгҖӮйҒ—дј жҖ§зјәд№ҸTlr5гҖҒIl10гҖҒTbx1гҖҒRag2зӯүе…Қз–«зӣёе…ізү©иҙЁзҡ„е°Ҹйј пјҢз”ұдәҺиӮ йҒ“иҸҢзҫӨеӨұи°ғпјҢжӣҙе®№жҳ“еҸ‘з”ҹз»“иӮ зӮҺеҸҠз»“иӮ зҷҢпјҢиҝҷз§Қзү№жҖ§е№¶дё”еҸҜд»Ҙдј йҖ’з»ҷеҒҘеә·е°Ҹйј гҖӮ
|
|
|
|
|
|
|
е…ұ1жқЎзІҫеҪ©еӣһеӨҚпјҢжңҖеҗҺеӣһеӨҚдәҺ 2016-11-21 13:45

е°ҡжңӘзӯҫеҲ°
4 иӮ йҒ“иҸҢзҫӨеңЁиӮҝзҳӨжІ»з–—дёӯдҪңз”Ё
иӮ йҒ“иҸҢзҫӨжңүзӣҠдәҺжҹҗдәӣиӮҝзҳӨжІ»з–—пјҢеңЁиӮҝзҳӨжң¬иә«жҲ–其并еҸ‘з—Үзҡ„жІ»з–—дёӯеҸ‘жҢҘйҮҚиҰҒдҪңз”ЁгҖӮз ”з©¶еҸ‘зҺ°пјҡ
иӮ йҒ“иҸҢзҫӨйҖҡиҝҮи°ғиҠӮиӮҝзҳӨеҫ®зҺҜеўғдёӯдёҚеҗҢй«“зі»з»Ҷиғһзҡ„еҠҹиғҪпјҢеҪұе“ҚиӮҝзҳӨе…Қз–«жІ»з–—еҸҠеҢ–з–—зҡ„еҸҚеә”гҖӮзҳӨеҶ…жіЁе°„ CpG еҜЎж ёиӢ·й…ёпјҲoligonucleotides, ODNпјүе…Қз–«жІ»з–—гҖҒиҒ”еҗҲIL-10дҝЎеҸ·жҠ—дҪ“дёӯе’ҢжІ»з–—еҸҜд»Ҙжңүж•ҲжІ»з–—жҷ®йҖҡе°Ҹйј зҡ„зҡ®дёӢжҺҘз§ҚиӮҝзҳӨпјҢдҪҶжҳҜеҜ№GFе°Ҹйј гҖҒжҠ—з”ҹзҙ жІ»з–—е°Ҹйј ж— ж•Ҳ[20]гҖӮеҜ№жҷ®йҖҡе°Ҹйј пјҢиҜҘз–—жі•еҸҜд»ҘеңЁж•°е°Ҹж—¶еҶ…еј•иө·TNFдҫқиө–жҖ§иӮҝзҳӨдёҘйҮҚеҮәиЎҖеқҸжӯ»пјҢиҜҘеҮәиЎҖеқҸжӯ»дҫқиө–иӮҝзҳӨзӣёе…іжҖ§еҲқе§Ӣй«“зі»з»Ҷиғһдә§з”ҹTNFеҸҠNOпјҢ继иҖҢCD40д»ӢеҜјDCжҝҖжҙ»гҖҒIL-12еҲҶжіҢгҖҒCD8+Tд»ӢеҜјзҡ„иӮҝзҳӨзү№ејӮжҖ§иҝҮ继е…Қз–«пјҢеҸ‘жҢҘжҢҒз»ӯзҡ„иӮҝзҳӨжқҖзҒӯдҪңз”ЁгҖӮдҪҶжҳҜпјҢеңЁиӮ йҒ“иҸҢзҫӨзјәд№Ҹе°Ҹйј пјҢCpG-ODNжІ»з–—еҗҺпјҢиӮҝзҳӨжөёж¶Ұй«“зі»з»Ҷиғһдә§з”ҹзӮҺз—Үз»Ҷиғһеӣ еӯҗеҰӮTNFгҖҒIL-12зҡ„иғҪеҠӣжҳҺжҳҫеүҠејұгҖӮеҸЈжңҚзҒҢжіЁLPS еҸҜд»ҘйғЁеҲҶжҒўеӨҚжҠ—з”ҹзҙ жІ»з–—е°Ҹйј еҜ№CpG-ODNжІ»з–—зҡ„еҸҚеә”дёҚиүҜгҖӮйҒ—дј жҖ§LPSеҸ—дҪ“зј–з Ғеҹәеӣ гҖҒ TLR4зј–з Ғеҹәеӣ зјәд№Ҹе°Ҹйј пјҢеҜ№CpG-ODNзҡ„еҸҚеә”иҰҒжҜ”йҮҺз”ҹеһӢе°Ҹйј дҪҺгҖӮжҸҗзӨәпјҡиӮ йҒ“иҸҢзҫӨLPSйҖҡиҝҮTLR4еҸ—дҪ“еҸҜд»ҘзӣҙжҺҘжҲ–й—ҙжҺҘжҝҖжҙ»иӮҝзҳӨжөёж¶Ұй«“зі»з»ҶиғһеҜ№TLR9й…ҚдҪ“CpG-ODNзҡ„еҸҚеә”гҖӮиӮ йҒ“иҸҢзҫӨз»„жҲҗдёҚеҗҢпјҢCpGOGNжІ»з–—еҗҺTNFзҡ„еҸҚеә”й«ҳдҪҺдёҚеҗҢгҖӮиӮ йҒ“иҸҢзҫӨдёӯжҹҗдәӣG+е’ҢG-зҡ„еӨҡе°‘дёҺиӮҝзҳӨй«“зі»з»ҶиғһеҜ№CpG-ODNеҸҚеә”е‘ҲжӯЈзӣёе…іпјҢиҖҢе…ұз”ҹд№іжқҶиҸҢж•°йҮҸеҲҷдёҺиӮҝзҳӨй«“зі»з»ҶиғһеҜ№ CpG-ODNеҸҚеә”е‘Ҳиҙҹзӣёе…ігҖӮG-Alistipes shaii еҸҜд»ҘеўһејәCpG-ODNзҡ„еҸҚеә”пјҢиҖҢд№іжқҶиҸҢL. fermentumеҲҷзӣёеҸҚеүҠејұCpG-ODNзҡ„еҸҚеә”[20]гҖӮ
й“Ӯзұ»еҢ–еҗҲзү©еҰӮиҚүй…ёй“ӮгҖҒйЎәй“Ӯзҡ„жІ»з–—ж•Ҳжһңд№ҹдёҺиӮ йҒ“иҸҢзҫӨеҜҶеҲҮзӣёе…і[20]гҖӮзҡ®дёӢ移жӨҚеҗҢж ·зҡ„иӮҝзҳӨпјҢжҠ—з”ҹзҙ жІ»з–—е°Ҹйј гҖҒGFе°Ҹйј еҜ№й“Ӯзұ»еҢ–еҗҲзү©еҰӮиҚүй…ёй“ӮгҖҒйЎәй“Ӯзҡ„еҸҚеә”жҳҫи‘—дҪҺдәҺжҷ®йҖҡе°Ҹйј гҖӮжҠ—з”ҹзҙ жІ»з–—в‘ жҠ‘еҲ¶иҚүй…ёй“ӮиҜұеҜјзҡ„зӮҺз—Үзӣёе…іеҹәеӣ гҖҒеҚ•ж ёз»ҶиғһеҲҶеҢ–гҖҒжҝҖжҙ»е’ҢеҠҹиғҪзӣёе…іеҹәеӣ зҡ„дёҠи°ғиЎЁиҫҫпјӣв‘ЎйҳІжӯўиҚүй…ёй“ӮиҜұеҜјзҡ„жӯЈеёёз»ҶиғһеҠҹиғҪеҰӮд»Ји°ўгҖҒиҪ¬еҪ•гҖҒзҝ»иҜ‘еҸҠDNAеӨҚеҲ¶зӣёе…іеҹәеӣ зҡ„дёӢи°ғиЎЁиҫҫпјӣв‘ўжҠ‘еҲ¶иӮҝзҳӨжөёж¶Ұй«“зі»з»ҶиғһйҖҡиҝҮNADPHж°§еҢ–й…¶NOX2зҡ„ROSдә§йҮҸпјҢд»ҺиҖҢеүҠејұй“Ӯзұ»еҢ–еҗҲзү©зҡ„жҠ—иӮҝзҳӨж•ҲжһңгҖӮдёҺCpG-ODNе…Қз–«жІ»з–—дёҚеҗҢпјҢиҚүй…ёй“Ӯзҡ„жІ»з–—ж•ҲжһңдёҚеҸ—TNFжҲ–TLR4еҪұе“ҚгҖӮ
з»Ҷиғһзү№ејӮжҜ’жҖ§CD8+Tз»Ҷиғһзҡ„иҝҮ继иҫ“жіЁжІ»з–—еҗҢж ·дёҺиӮ йҒ“иҸҢзҫӨжңүе…ігҖӮе…Ёиә«з…§е°„пјҲtotal body irradiation, TBIпјүжІ»з–—еҸҜд»ҘжҸҗй«ҳCD8+Tз»ҶиғһиҝҮ继е…Қз–«жІ»з–—зҡ„ж•ҲжһңпјҢжңүзӣҠдәҺжҝҖжҙ»DCпјҢжңүзӣҠдәҺеҶ…зҺҜеўғзЁіе®ҡз»Ҷиғһеӣ еӯҗзҡ„дә§з”ҹгҖӮTBIдҝғиҝӣзҡ„иӮ зі»иҶңж·Ӣе·ҙз»“з»ҶиҸҢ移дҪҚгҖҒеҚҮй«ҳдәҶиЎҖжөҶLPSж°ҙе№ігҖӮжҠ—з”ҹзҙ жІ»з–—гҖҒеӨҡй»ҸиҸҢзҙ Bдёӯе’ҢиЎҖжөҶLPSгҖҒйҳ»ж–ӯLPSдҝЎеҸ·дј еҜјйғҪеҸҜд»ҘеүҠејұTBIеҸҠCD8+Tз»ҶиғһиҝҮ继е…Қз–«жІ»з–—зҡ„ж•ҲжһңгҖӮз»ҷжңӘз…§е°„е°Ҹйј иҫ“жіЁLPSпјҢеҸҜд»ҘеўһеҠ иҝҮ继иҫ“жіЁCD8+Tз»Ҷиғһзҡ„ж•°йҮҸпјҢеўһејәе…¶еҠҹиғҪпјҢеёҰжқҘиӮҝзҳӨзҡ„й•ҝжңҹжІ»ж„Ҳ[21]гҖӮдёҺиҚүй…ёй“Ӯзұ»дјјпјҢзҺҜзЈ·й…°иғәпјҲcyclophosphamide, CTXпјүйҷӨзӣҙжҺҘз»ҶиғһжҜ’жҖ§еӨ–пјҢиҝҳиҜұеҜје…Қз–«еҺҹжҖ§з»Ҷиғһжӯ»дәЎпјҲimmunogenic cell deathпјүпјҢиҝӣиҖҢеј•еҮәиҮӘйҖӮ
еә”жҠ—иӮҝзҳӨе…Қз–«еҸҚеә”пјҲadaptive antitumor immune responseпјүпјҢеҗҢж—¶дә§з”ҹиӮҝзҳӨзү№ејӮжҖ§CTLsгҖӮCTX жқҖдјӨиӮҝзҳӨгҖҒиҜұеҜје…Қз–«еҸҚеә”зҡ„дҪңз”Ёдҫқиө–дәҺиӮ йҒ“иҸҢзҫӨпјҢGFе°Ҹйј жҲ–жҠ—з”ҹзҙ жІ»з–—е°Ҹйј еҜ№CTXдёҚж•Ҹж„ҹгҖӮеҜ№жҷ®йҖҡе°Ҹйј пјҢCTXжІ»з–—ж”№еҸҳдәҶиӮ йҒ“иҸҢзҫӨзҡ„з»„жҲҗпјҢиҜұеҸ‘й»ҸиҶңзӮҺеҸҠеј•жөҒж·Ӣе·ҙз»“зҡ„G+з»ҶиҸҢ移дҪҚпјҢеўһејәж•Ҳеә”Th17з»ҶиғһеҸҠи®°еҝҶTh1з»Ҷиғһе…Қз–«еҸҚеә”[22]гҖӮеҢ–з–—иҚҜзү©еј•иө·зҡ„е…Қз–«еҺҹжҖ§з»Ҷиғһжӯ»дәЎжҝҖжҙ» APCsгҖҒиҜұеҜјжҠ—иӮҝзҳӨе…Қз–«еҸҚеә”пјҢиҝҷдёӘдҪңз”ЁдёҚд»…дёҺеқҸжӯ»з»„з»ҮйҮҠж”ҫзҡ„зӮҺз—Үд»ӢиҙЁжңүе…іпјҢиҝҳдёҺиӮ йҒ“иҸҢзҫӨеҸҠе…¶дә§зү©зҡ„йў„жҝҖжҲ–еўһејәжңүе…іпјҢи§Ғеӣҫ4гҖӮ
еӣҫзүҮиҜ·зӮ№еҮ»дёҠж–№й“ҫжҺҘ
з»јдёҠжүҖиҝ°пјҢжҲ‘们еҸҜд»Ҙжё…жҘҡең°зңӢеҲ°пјҢиӮ йҒ“иҸҢзҫӨеҸҜд»ҘеңЁеӨҡдёӘзҺҜиҠӮдёҠеҪұе“ҚиӮҝзҳӨзҡ„еҸ‘з”ҹгҖҒеҸ‘еұ•дёҺжІ»з–—гҖӮ
йүҙдәҺиӮ йҒ“иҸҢзҫӨеңЁиӮҝзҳӨеҸ‘з”ҹгҖҒеҸ‘еұ•еҸҠжІ»з–—дёӯзҡ„йҮҚиҰҒдҪңз”ЁпјҢжҲ‘们жңүзҗҶз”ұзӣёдҝЎпјҡдёҚд№…зҡ„е°ҶжқҘпјҢзІӘиҸҢ移жӨҚе°Ҷдјҡиў«з”ЁдәҺиӮҝзҳӨжІ»з–—пјҢд»ҺиҖҢдә§з”ҹдёҖдёӘеҙӯж–°зҡ„иӮҝзҳӨжІ»з–—ж–№жі•вҖ”иӮҝзҳӨзІӘиҸҢз–—жі•пјҲcancer?fecal?microbiota?therapy, CFMTпјүгҖӮCFMTд»ҺжҠҖжңҜдёҠе·Із»Ҹе®Ңе…ЁжҲҗзҶҹпјҢеҹәжң¬жҖқи·ҜжңүдәҢпјҡв‘ еҜ№иӮҝзҳӨжӮЈиҖ…зҡ„иӮ йҒ“иҸҢзҫӨиҝӣиЎҢеҢ–еӯҰе’ҢпјҲжҲ–пјүзү©зҗҶжё…йҷӨеҗҺпјҢе°ҶеҒҘеә·дҫӣиҖ…зҡ„зІӘиҸҢжҲ–иҖ…дәәе·Ҙж”№йҖ зҡ„зІӘиҸҢ移жӨҚз»ҷиӮҝзҳӨжӮЈиҖ…пјӣв‘Ўе°ҶиӮҝзҳӨжӮЈиҖ…зІӘиҸҢеҸ–еҮәеҗҺпјҢеңЁдҪ“еӨ–иҝӣиЎҢдәәе·Ҙж”№йҖ пјҢ然еҗҺеӣһиҫ“з»ҷиӮҝзҳӨжӮЈиҖ…гҖӮдҪҶжҳҜпјҢе®һйҷ…еә”з”Ёж–№йқўд»Қ然жңүеҫҲиҝңзҡ„и·ҜиҰҒиө°гҖӮиӮ йҒ“иҸҢзҫӨдёҺиӮҝзҳӨзҡ„е…ізі»жҳҜиҝ‘е№ҙжқҘиӮҝзҳӨз ”з©¶зҡ„дёҖдёӘзғӯзӮ№йўҶеҹҹгҖӮй«ҳйҖҡйҮҸеҫ®з”ҹзү©и¶…еҸҳ16Sж ёзі–дҪ“RNAеҹәеӣ жөӢеәҸеҸҠз”ҹзү©дҝЎжҒҜеӯҰз®—жі•пјҲbioinformatic algorithmsпјүзҡ„дҪҝз”ЁпјҢдҪҝз ”з©¶дәәе‘ҳеҸҜд»ҘйӣҶзҫӨеҲҶжһҗжҹҗиӮ йҒ“еҫ®з”ҹзү©зҫӨеңЁиӮҝзҳӨеҸҠеҶ…зҺҜеўғзЁіе®ҡдёӯзҡ„з»јеҗҲдҪңз”ЁпјҢиҖҢж— йңҖиҝӣиЎҢеҚ•дёӘеҲҶзҰ»еҸҠеҹ№е…»еҲҶжһҗгҖӮжҠҖжңҜзҡ„еҲӣж–°дҪҝз ”з©¶дәәе‘ҳеҫҲе®№жҳ“еҲҶиҫЁеҮәиҮҙзҷҢз»ҶиҸҢпјҲеҰӮ Escherichia coli, Fusobacterium nucleatum, Bacteroides fragilisпјүе’ҢжҠ—зҷҢз»ҶиҸҢ [еҰӮEubacterium, Lactobacillus, Bacteriodes, ClostridiumпјҲXIVa and IVaпјү, Faecalibacterium, Bifdobacterium, Roseburia]пјҢеҫҲе®№жҳ“дәҶи§ЈиӮҝзҳӨжӮЈиҖ…иӮ йҒ“иҸҢзҫӨзҡ„еҫ®з”ҹзү©жЁЎејҸпјҲmicrobial profleпјүпјҢиҝӣиҖҢй’ҲеҜ№жҖ§дәәе·Ҙж”№йҖ иӮ йҒ“иҸҢзҫӨпјҢд»ҺиҖҢдёәиӮҝзҳӨйў„йҳІгҖҒиӮҝзҳӨжІ»з–—жҸҗдҫӣдәҶеҸҜиғҪгҖӮ |
|
|
|
|
|
|



 дҫқжІғиҘҝз”ЁиҚҜеҗҺпјҢиҜҘжҖҺд№ҲеҠһпјҹ
дҫқжІғиҘҝз”ЁиҚҜеҗҺпјҢиҜҘжҖҺд№ҲеҠһпјҹ
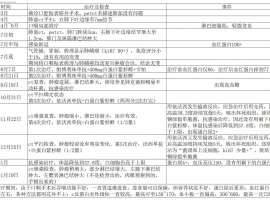
жҲ‘зҲёзҲёдәҺ2024е№ҙ7жңҲзЎ®иҜҠиӮәйіһзҷҢпјҢж•ҙдёӘжІ»з–—иҝҮзЁӢеҰӮйҷ„件йҮҢиЎЁж ј
дҫқжІғиҘҝз”ЁиҚҜеҗҺпјҢиҜҘжҖҺд№ҲеҠһпјҹ
дҫқжІғиҘҝз”ЁиҚҜеҗҺпјҢиҜҘжҖҺд№ҲеҠһпјҹ
жҲ‘зҲёзҲёдәҺ2024е№ҙ7жңҲзЎ®иҜҠиӮәйіһзҷҢпјҢж•ҙдёӘжІ»з–—иҝҮзЁӢеҰӮйҷ„件йҮҢиЎЁж ј
 и„‘иҶңиҪ¬з§»иҜҠз–—жҪҳжҢҜе®Үж•ҷжҺҲ科жҷ®жҖ»з»“
и„‘иҶңиҪ¬з§»иҜҠз–—жҪҳжҢҜе®Үж•ҷжҺҲ科жҷ®жҖ»з»“
и„‘иҶңиҪ¬з§» дёӯдҪҚжІ»з–—з”ҹеӯҳжңҹ 3-6жңҲ
и„‘иҶңиҪ¬з§»зҡ„дёҙеәҠиЎЁзҺ°
и„‘иҶңиҪ¬з§»иҜҠз–—жҪҳжҢҜе®Үж•ҷжҺҲ科жҷ®жҖ»з»“
и„‘иҶңиҪ¬з§»иҜҠз–—жҪҳжҢҜе®Үж•ҷжҺҲ科жҷ®жҖ»з»“
и„‘иҶңиҪ¬з§» дёӯдҪҚжІ»з–—з”ҹеӯҳжңҹ 3-6жңҲ
и„‘иҶңиҪ¬з§»зҡ„дёҙеәҠиЎЁзҺ°
 йғҪиҜҙе…Қз–«дёҚе…Ҙи„‘пјҢе…ҘдәҶе°ұдёҚеҫ—дәҶ
е…Қз–«жІ»з–—д»ҘеҗҺзҡ„з–—ж•ҲеҸҚеә”пјҢеҪ“з—…зҒ¶еўһеӨ§ж—¶пјҢдёҚдёҖе®ҡжҳҜиҝӣеұ•пјҢиҝҳжңүдёҖз§ҚеҸҜиғҪпјҢеҸ«еҒҡCRпјҢиҝҷдёӘзҺ°
йғҪиҜҙе…Қз–«дёҚе…Ҙи„‘пјҢе…ҘдәҶе°ұдёҚеҫ—дәҶ
е…Қз–«жІ»з–—д»ҘеҗҺзҡ„з–—ж•ҲеҸҚеә”пјҢеҪ“з—…зҒ¶еўһеӨ§ж—¶пјҢдёҚдёҖе®ҡжҳҜиҝӣеұ•пјҢиҝҳжңүдёҖз§ҚеҸҜиғҪпјҢеҸ«еҒҡCRпјҢиҝҷдёӘзҺ°


 жҸҗеҚҮеҚЎ
жҸҗеҚҮеҚЎ зҪ®йЎ¶еҚЎ
зҪ®йЎ¶еҚЎ жІүй»ҳеҚЎ
жІүй»ҳеҚЎ е–§еҡЈеҚЎ
е–§еҡЈеҚЎ еҸҳиүІеҚЎ
еҸҳиүІеҚЎ еҚғж–ӨйЎ¶
еҚғж–ӨйЎ¶ жҳҫиә«еҚЎ
жҳҫиә«еҚЎ